 Setting up apparatus
Setting up apparatus Video capture & tracking
Video capture & tracking Observing behaviour
Observing behaviour Connecting equipment
Connecting equipment Automating complex tests
Automating complex tests Running tests
Running tests Results
Results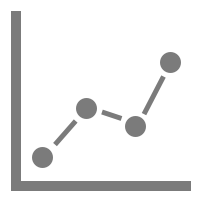 Visualising data
Visualising data Analysis
Analysis Transferring data
Transferring data Open field
Open field Water-maze
Water-maze Y-maze
Y-maze Fear conditioning
Fear conditioning Novel object
Novel object Barnes maze
Barnes maze Radial arm maze
Radial arm maze Light/dark box
Light/dark box Operant conditioning
Operant conditioning Zebrafish
Zebrafish Computers
Computers Multifunction remote
Multifunction remote Accessories
Accessories Digital interface
Digital interface Optogenetic interface
Optogenetic interface Synchronisation interface
Synchronisation interface Relay interface
Relay interface Audio interface
Audio interface Touch interface
Touch interface Analogue interface
Analogue interface USB TTL cable
USB TTL cable Animal shocker
Animal shocker Components
Components Place preference
Place preference ANY-box
ANY-box T-maze
T-maze Zero maze
Zero maze Hole board
Hole board Sociability cage
Sociability cage OPAD
OPAD RAPC
RAPC Waterwheel forced swim test
Waterwheel forced swim test Thermal gradient ring
Thermal gradient ring Operon
Operon Activity Wheel
Activity Wheel Full ANY-maze licence
Full ANY-maze licence Other licence types
Other licence types Developing countries licence
Developing countries licence Contact support
Contact support Support Policy
Support Policy FAQs
FAQs Guides
Guides Downloads
Downloads Send us files
Send us files Activate a licence ID
Activate a licence ID Documentation
Documentation Contact us
Contact us Blog
Blog About
About Testimonials
Testimonials Privacy Policy
Privacy Policy
The key to automating the elevated plus-maze is to detect zone entries and exits with precision
ANY-maze’s whole body tracking provides this feature, which together with flexible results analysis, make ANY-maze an ideal system for plus-maze work - see the Benefits tab, below, to learn more.
On the other tabs you'll find some videos of real plus-maze tests, as well as details of recommended equipment and a list of results that are especially useful in this test.

Precision tracking
ANY-maze’s whole body tracking allows you to specify precisely when an arm entry should be scored.
For example, specifying an open-arm entry as occurring when 80% of the animal’s body is in the arm, equates very well with the traditional four-paws-in-the-arm rule.
Watch the video on the right to see this in action.


What to do about the centre zone?
Exactly how entries into the centre zone of the plus-maze should be scored is a source of some controversy. ANY-maze doesn’t force you to score entries in any particular way – rather it allows you to define exactly when the system should consider the animal is the centre.

Flexible results
Another area where opinions differ about the plus-maze test is precisely how to calculate the percentage of time in the open (or closed) arms.
Some researchers choose to include the time in the centre in this calculation, while others ignore the centre.
In ANY-maze, you can define this measure in whatever way you prefer (the image on the right shows one way to calculate it), and if you change your mind ANY-maze will immediately re-calculate the results accordingly.

Getting more from your data
In ANY-maze you can alter the definition of your zones (or indeed almost anything else) at any time, whether before, during or after you’ve run your tests.
For example, some researchers report the amount of time the animal spends on the ends of the open arms. If you ran a plus-maze experiment without including this zone, you could simply add it afterwards.

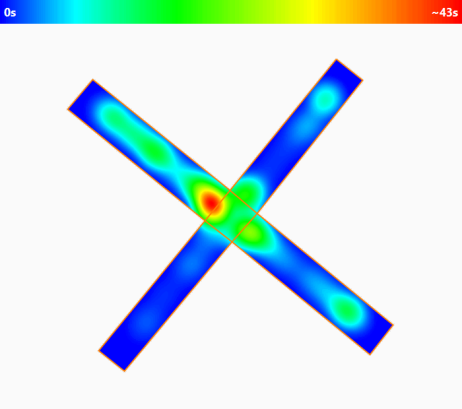
Viewing the animal's track
ANY-maze can plot the animal’s track as a line or as a heat map.
Heat maps, indicate how much time the animal spent in different parts of the apparatus and can either show data for individual tests (as shown here) or averaged data for different groups.
In this example, we can see that the animal spent most time at the exit of the closed arms.
Results
ANY-maze can provide literally hundreds of results for any test, but some of those that are commonly used in the plus-maze include:
- Time in the open arms
- Time in the closed arms
- Time in the centre arms
- Number of entries into the open arms
- Number of entries into the closed arms
- Total distance travelled
- Percentage of time in the open arms

Tracking a dark mouse in an unevenly
lit plus-maze
In this plus-maze there is very low contrast between the animal and the background of the closed arms. Despite this (and the overall poor quality of the recording) ANY-maze still tracks reliably and accurately detects the animal’s entries and exits to the arms using whole body tracking – which as can be seen, approximates well to the four-paws-in-the-arm rule.

Tracking a white rat with a dark skull cap in a white plus-maze
In this example a white rat, which has been fitted with a dark skull cap, is tracked in a white plus-maze. The skull cap provides a much higher contrast target than the animal, but ANY-maze correctly tracks the entire animal and accurately uses whole body tracking to detect arm entries – once again closely mirroring the traditional four-paws-in-the-arm rule.

Mazes
We manufacture our own plus-mazes, both for Rat and Mouse. The walls of the closed arms simply lift off for easy cleaning and the entire maze is mounted on detachable legs which provide convenience for storage.
View more
Cameras
USB camera
The ANY-maze USB camera is an excellent choice for the plus-maze. We recommend fitting this camera with a varifocal (zoom) lens, so you can simply mount the camera on the ceiling and then zoom in and out until the maze nicely fits the camera's view.
View more
Web cam
A webcam is usually a good, and inexpensive, alternative choice for the plus-maze. If you intend to test in normal lighting conditions (>= 100 lux) and you can mount the camera far enough from the maze for it to see it all, then a webcam should work well.

Lighting
Infrared illuminator
If you intend to track in low light (< 10 lux) or in darkness, then you may wish to use an infrared illuminator and an infrared sensitive camera (most cameras are IR sensitive).
View more
Accessories
ANY‑maze Radio remote control
The ANY‑maze Radio remote control provides a convenient way to start the test as soon as the animal is in the maze. This remote works through walls and has 2 buttons, allowing you to control two apparatus independently.
View more
Achilly, N.P., Wang, W. & Zoghbi, H.Y. (2021) Presymptomatic training mitigates functional deficits in a mouse model of Rett syndrome. Nature 592, 596–600
Mungenast, H.K., et al (2021) Impact of Light–Dark Phase on Delay Discounting Behavior in Rats. Psychol Rec. https://doi.org/10.1007/s40732-021-00462-1
Tan, S.Z.K., et al (2021) Prelimbic Cortical Stimulation Disrupts Fear Memory Consolidation through Ventral Hippocampal Dopamine 2 Receptors. British Journal of Pharmacology https://doi.org/10.1111/bph.15505
Fyke, W., et al (2021) Pharmacological inhibition of the primary endocannabinoid producing enzyme, DGL‐α, induces autism spectrum disorder‐like and co‐morbid phenotypes in adult C57BL/J mice. Autism Research https://doi.org/10.1002/aur.2520
Assali, A., et al (2021) Sex-dependent role for EPHB2 in brain development and autism-associated behavior. Neuropsychopharmacology https://doi.org/10.1038/s41386-021-00986-8
Diniz, C.V.P., et al (2021) Classical Music and Environmental Enrichment Enhanced Spatial Memory and Learning and Increased Mouse Innate Tendency to Avoid Open Spaces. EC Neurology 13.1
Hao, T., et al (2020) Astrocytes-induced neuronal inhibition contributes to depressive-like behaviors during chronic stress. Life Sciences 258, 118099
Barros-Santos, T., et al (2020) Effects of chronic treatment with new strains of Lactobacillus plantarum on cognitive, anxiety- and depressive-like behaviors in male mice. PloS one, https://doi.org/10.1371/journal.pone.0234037
Abela, A.R., et al (2020) Median raphe serotonin neurons promote anxiety-like behavior via inputs to the dorsal hippocampus. Neuropharmacology 168, 107985
Wiersielis, K.R., et al (2020) Maternal exposure to organophosphate flame retardants alters locomotor and anxiety-like behavior in male and female adult offspring. Hormones and Behavior, 122, 104759
Davis K.C., et al (2020) Behavioral Alterations in Mice Carrying Homozygous HDAC4A778T Missense Mutation Associated With Eating Disorder. Frontiers in Neuroscience https://doi.org/10.3389/fnins.2020.00139
Ke, X., et al (2020) Adverse maternal environment and western diet impairs cognitive function and alters hippocampal glucocorticoid receptor promoter methylation in male mice. Physiological Reports, 8(8), e14407
Terstege, D.J., et al (2020) Standardised ginseng extract G115® potentiates the antidepressant-like properties of fluoxetine in the forced swim test. Acta Neuropsychiatrica, 33(3), 141, doi:10.1017/neu.2021.2
J.V. Cabrel-Costa et al. (2018) Intermittent fasting uncovers and rescues cognitive phenotypes in PTEN neuronal haploinsufficient mice; Scientific Reports 2018; 8:8595
L.R. Meyer et al. (2018) Perinatal SSRI exposure permanently alters cerebral serotonin receptor mRNA in mice but does not impact adult behaviors; The Journal of Maternal-Fetal & Neonatal Medicine 2018; 31(11): 1393-1401
R.D. Brose et al. (2018) Hydroxyurea attenuates oxidative, metabolic, and excitotoxic stress in rat hippocampal neurons and improves spatial memory in a mouse model of Alzheimer’s disease; Neurobiology of Aging 2018; 72:121-133
R.A. Déziel & R.A. Tasker (2018) Bilateral ischaemic lesions of the medial prefrontal cortex are anxiogenic in the rat; Acta Neuropsychiatrica 2018; 30(3); 181-186
A.L. Russel et al. (2018) Sex-Dependent Effects of Mild Blast-induced Traumatic Brain Injury on Corticotropin-releasing Factor Receptor Gene Expression: Potential Link to Anxiety-like Behaviors; Neuroscience 2018; 392:1-12
Somkuwar, Sucharita S., et al. (2017) Abstinence from prolonged ethanol exposure affects plasma corticosterone, glucocorticoid receptor signaling and stress-related behaviors. Psychoneuroendocrinology (2017).
Laaksonen, Sakari, et al. (2017) Behaviour, stress and welfare of Sprague Dawley rats (Rattus norvegicus) on diet board feeding for 24 months. Applied Animal Behaviour Science (2017).
Karl F, Grießhammer A, Üçeyler N and Sommer C (2017) Differential Impact of miR-21 on Pain and Associated Affective and Cognitive Behavior after Spared Nerve Injury in B7-H1 ko Mouse. Front. Mol. Neurosci. 10:219.
Y Ye et al. (2017) Alterations in Opioid Inhibition Cause Widespread Nociception but Do Not Affect Anxiety-Like Behavior in Oral Cancer Mice Neuroscience. S0306-4522(17)30443-8
K Shang et al. (2017) Parent-of-origin effects on schizophrenia-relevant behaviours of type III neuregulin 1 mutant mice, Behav Brain Res 2017;332:250-258.
B Riaz et al. (2017) Oral supplementation of Thuja oreintalis leaf extract moderately affects the behavior in albino mice in a gender specific manner. Pak J Pharm Sci 2017;30(3):683-688
Vargish, G. A., et al. (2016) Persistent inhibitory circuit defects and disrupted social behaviour following in utero exogenous cannabinoid exposure. Molecular psychiatry (2016).
Leão, Anderson HFF, et al. (2016) Hippocampal-dependent memory in the plus-maze discriminative avoidance task: The role of spatial cues and CA1 activity. Behavioural brain research 304 (2016): 24-33.
Hermes, Gretchen, et al. (2016) Role of mitochondrial uncoupling protein-2 (UCP2) in higher brain functions, neuronal plasticity and network oscillation. Molecular Metabolism (2016).
Briassoulis, George, et al. (2016) Studies of mice with cyclic AMP-dependent protein kinase (PKA) defects reveal the critical role of PKA’s catalytic subunits in anxiety. Behavioural Brain Research 307 (2016): 1-10.
Singh, Prabhakar, et al. (2016) GRK5 deficiency leads to susceptibility to intermittent hypoxia-induced cognitive impairment. Behavioural brain research (2016).